کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)

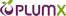
32,400 تومانشناسه فایل: 6204
چکیده
نورون های آینه ای برای افروختن در دو حالت فعالیتی را انجام می دهند و یکی زمانی که حیوانی شیء را به شیوه ای خاص دستکاری می کند و زمانی که حیوانی دیگر (یا آزمایشگر) را می بیند که کم و بیش به یکدیگر شباهت دارد. این نورون ها در ابتدا در میمون های ماکاک، در قشر مخ پیش حرکتی پیشین، ناحیه F5 و بعد از آن نیز قسمت لگن پاریتال (آهیانه) یافت شده بودند. داده های عصبی جدید نشان می دهد که مغز انسان بالغ با یک سیستم نورون آینه ای، که شامل نورون های آینه ای و نورون های دیگر به منظور تطبیق مشاهده و اجرای فعالیت ها است. نورون های آینه ای برای تشخیص فعالیت در میمون ها و نیز انسانها به کار می روند، در حالی که نقش احتمالی آن ها در تقلید و زبان است که می تواند در انسان ها و نه میمون ها تحقق یابد. این مقاله، نقش مهم مدل های محاسباتی در ارائه توضیحات کافی و علیت برای پدیده های مشاهده شده شامل سیستم های آینه ای و فرآیندهای یادگیری که آن ها را تشکیل می دهد، نشان می دهد و نیاز به حوزه های بیشتر برای پیشرفت مدار نورون آینه ای میمون را برای شناخت فرضی نسبت داده شده به سیستم های نورون آینه ای انسان را تقویت نماید.
مقدمه مقاله
نورون های آینه ای برای افروختن در دو حالت فعالیتی را انجام می دهند و یکی زمانی که حیوانی شیء را به شیوه ای خاص دستکاری می کند و زمانی که حیوانی دیگر (یا آزمایشگر) را می بیند که کم و بیش به یکدیگر شباهت دارد. این نورون ها در ابتدا در میمون های ماکاک، در قشر مخ پیش حرکتی پیشین، ناحیه F5 [21,32,76] و سپس در قسمت کوچک آهیانه ای تحتانی [28،78] یافت شد. ویژگی آینه ای در تمام نورون های دیداری حرکتی F5 مشاهده نشده است. نورون های دیداری حرکتی F5 وجود دارند که به صورت انتخابی برای ارائه تصویری شیء داده شده تخلیه میشود که همچنین در هنگام در نظر گرفتن آن مفهوم نیز به صورت گزینشی آزاد می شود[61]. این نورون ها موسوم به نورون های معیار هستند و اعتقاد بر این است که نقش مهمی در تبدیل ظاهر دیداری اشیاء به طرح های حرکتی به منظور تعامل با آن ها است [77]. منطقه F5 نیز مشتمل بر نورون های آینه ای شنوایی [54] است که نه تنها به آن منظره پاسخ میدهد بلکه به صدای آن فعالیت ها با اصوات معمولی (مثل شکستن بادام زمینی، پاره کردن کاغذ) پاسخ می دهد. اگرچه به طور کلاسیک این اقدامات به نورون های آینه ای میمون هایی که متغیر در نظر گرفته می شوند، ارتباط دارد، یعنی این فعالیت باید به یک شیء توجه داشته باشد، که ممکن است پیش از تماس دستی از مقابل دیدگاه میمون حذف شود [به عنوان مثال، 95]، بیشتر داده های جدید نشان می دهد که انتقال پذیری ممکن است یک پیش شرط برای فعالیت شبه آینه ای نباشد (همچنین به بخش 7 مراجعه کنید).
ABSTRACT Mirror neurons Functions, mechanisms and models
Mirror neurons for manipulation fire both when the animal manipulates an object in a specific way and when it sees another animal (or the experimenter) perform an action that is more or less similar. Such neurons were originally found in macaque monkeys, in the ventral premotor cortex, area F5 and later also in the inferior parietal lobule. Recent neuroimaging data indicate that the adult human brain is endowed with a “mirror neuron system,” putatively containing mirror neurons and other neurons, for matching the observation and execution of actions. Mirror neurons may serve action recognition in monkeys as well as humans, whereas their putative role in imitation and language may be realized in human but not in monkey. This article shows the important role of computational models in providing sufficient and causal explanations for the observed phenomena involving mirror systems and the learning processes which form them, and underlines the need for additional circuitry to lift up the monkey mirror neuron circuit to sustain the posited cognitive functions attributed to the human mirror neuron system.
Introduction
Mirror neurons for manipulation fire both when the animal manipulates an object in a specific way and when it sees another animal (or the experimenter) perform an action that is more or less similar. Such neurons were originally found in macaque monkeys, in the ventral premotor cortex, area F5 [21,32,76], and later also in the inferior parietal lobule [28,78]. Not all visuomotor neurons in F5 show the mirror property. There are F5 visuomotor neurons that selectively discharge to the visual presentation of a given object, which also discharge selectively during grasping of that object [61]. These neurons are called canonical neurons, and are believed to play a crucial role in transforming visual appearance of objects into motor plans for interacting with them [77]. Area F5 also includes auditory mirror neurons [54] that respond not only to the view but also to the sound of actions with typical sounds (e.g. breaking a peanut, tearing paper). Although classically, the actions associated with mirror neurons in the monkey are considered to be transitive, i.e. the action has to be directed to an object, which may be removed from the view of the monkey before hand-contact [e.g. 95], more recent data indicates that transitivity may not be a prerequisite for mirror-like activity (see also Section 7).
Recent neuroimaging findings indicate that the adult human brain is endowed with a system for matching the observation and execution of actions which might be homologous to the macaque mirror neuron system [10,73,77]. In spite of the growing number of human brain imaging data related to posited ‘human mirror systems’ [e.g. 6,13,25,43,44,83], the experimental data on mirror neurons are available mainly for monkeys as systematic recordings using electrophysiology cannot be used to investigate the human brain. Therefore it must be emphasized that in humans a mirror system refers to a brain region (or set of brain regions) that becomes active for both observation and execution of a class of actions.
In the literature a set of functions is attributed to monkey and human putative mirror neuron system, and several terms are used to describe mechanisms underlying these functions. However, many of these functions are observed in human but not in monkeys, thus suggesting evolution within the mirror systems or within the wider networks of which they are part. Among these are imitation [e.g. 13,58], action understanding [e.g. 95], intention attribution [43] and (evolution of) language [74]. Recently, reviews and metaanalyses that are critical of the claimed mirror neuron functions have started to appear, in particular with the focus on the ambiguity of the terminology used to describe mirror neuron functions such as direct matching and motor resonance [17,19,23,92]. In part such failures to unambiguously describe mirror neuron function, follow from ignoring the distinction between a brain function and a possible mechanism (Box 1). In the majority of mirror neuron literature, functions associated with a mirror system in humans are attributed to “direct matching” or “motor resonance” and sometimes with “motor simulation” as a mechanism to underlie action/intention understanding [34,78] and theory of mind [33] without either a precise definition of such a “mechanism” nor a clear account of how it contributes to the observed function. It is simply assumed that mirror systems are involved in this property. However, we know (by extrapolation from the macaque) that a mirror system will contain many types of neurons other than mirror neurons. Thus, when a brain imaging study reports increased activity in a mirror system for some task relative to a control it is a mistake (all too frequent in the literature) to assume that activity in mirror neurons underlies the activation – in some cases, it will be, but by no means in all.
Our task in this article is to make the case for the use of computational models, whether in terms of neural networks or higher level constructs such as control systems, in clarifying mechanisms that are sufficient to explain observed phenomena involving mirror systems. The mechanisms we demonstrate might not be the same with those employed by the brain, but their very precision sets the stage for experiments more precise than those guided by claims like “a motor resonance supports the observed function.” To be concrete, we will be interested with these questions:
- Is the postulated mechanism sound from a computational point of view?
- To what extent can a brain implement the postulated mechanism?
- What are the additional mechanisms needed on top of the claimed mechanisms to yield the posited functions?
![نمایش استفاده از نورون های آینهای در الگوی مفهومی گلدمن و گالیس [33] به منظور درک مقصود](https://maghalejoo.com/wp-content/uploads/2019/02/02/img-2.jpg)
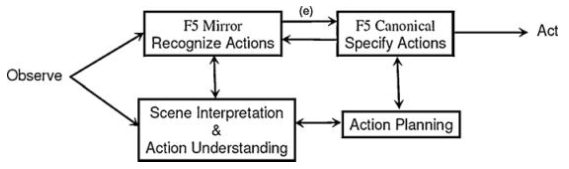
![نمایش درک مقصود و پیشبینی حسی رو به جلو در رابطه با نقش MN ها در الگوی محاسباتی اوزتوپ و همکاران [69].](https://maghalejoo.com/wp-content/uploads/2019/02/02/img-3.jpg)
- مقاله درمورد کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- نورون های آینه: توابع، مکانیزم ها و مدل ها
- کارآیی ها، ساز و کارها و الگوهای نورون های آینه ای
- پروژه دانشجویی کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- عملکرد ها و مکانیزم نورون های آینه ای
- پایان نامه در مورد کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- تحقیق درباره کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- مقاله دانشجویی کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN) در قالب پاياننامه
- پروپوزال در مورد کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- گزارش سمینار در مورد کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)
- گزارش کارورزی درباره کارکرد، مکانیزم و الگوهای نورون های آینه ای (MN)