نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
شاپا پرینت: 0022-0795 وبسایت مرجع 122 رفرنس دارد
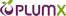
29,500 تومانشناسه فایل: 9703
مقدمه مقاله
به خوبی ثابت شده است که بیماران مبتلا به بیماریهای متابولیکی، به ویژه دیابت مقاوم به انسولین و دیابت ملیتوس نوع 2 (T2DM) بیش تر از دو برابر احتمال ایجاد بیماری قلبی عروقی تسریع یافته (CVD) مانند آترواسکلروز، سکته و بیماری عروق کرونر است (reviewed in Rask-Madsen & King (2013)). بیماری عروق کرونر علت اصلی مرگ در بیماران T2DM است و خطر مرگ و میر در زمان در مقایسه با مردان بسیار بیشتر است (Peters et al. 2014).
بیماری شدید عروق کرونر میتواند نتیجهای از انفارکتوس قلبی و کاهش شدید عملکرد قلب شده و منجر به ایجاد نارسایی قلبی خواهد شد (Hausenloy & Yellon 2013). خوشهای از فاکتورهای خطر اخیرا بوسیلهی انجمن دیابت آمریکا و کالج آمریکایی بنیاد قلب و عروق به عنوان شاخصهای قابل اعتماد خطر ابتلا به T2DM و CVD تحت عنوان خطر متابولیسمی قلبی عروقی تعریف شدهاند (CMR; Brunzell et al. 2008). این خطرات عبارتند از چاقی، قند خون بالا، فشارخون بالا، مقاومت به انسولین و دیسپیدمی. حضور CVD ثانویه در بیماران مبتلا به ایسکمی-پرفیوژن مجدد (IR) یا T2DM میتواند با عنوان بیماریهای متابولیسمی– قلبی عروقی (CMD) مورد بررسی قرار گیرد. با توجه به شیوع روزافزون و عواقب شدید CMD، رویکردهای جدیدی برای تشخیص و درمان CMD لازم هستند. وزیکولهای خارج سلولی (EVs) وزیکولهایی کوچک (50 نانومتر تا 2 میلیمتر) هستند که از سطح انواع مختلفی از سلولها به مایعات بدنی مانند پلاسما، شیر، بزاق، عرق، اشک، منی و ادرار ترشح میشوند. کلاسهای مختلفی از وزیکولهای خارج سلولی وجود دارند که عبارتند از اگزوزومها، میکرووزیکولهای (MVs) و اجسام آپوپتوزی که توسط مکانیسمهای مختلفی ساخته میشوند. اگزوزومها احتمالا بیشترین توجهات را به خود جلب کردهاند، اینها جمعیتی همگن از وزیکولهای خارج سلولی هستند که در زمان اتصال اجسام چند وزیکولی (MVB؛ گاهی با نام اندوزومهای چند وزیکولی یا MVE نیز نامیده میشوند) به غشاء پلاسمایی در فرایندهای شدیدا حفاظت شده منتشر شده و محتویاتشان را رها میکنند. سلولها میتوانند جمعیت ناهمگنتری از وزیکولهای خارج سلولی با قطر بیش از 2 میلیمتر و با نام میکرووزیکولها را تولید کنند که با جوانهزنی و بیرون ریزی غشاء پلاسمایی تشکیل میشوند، این فرایند شامل مسیرهای پیامرسانی وابسته به کلسیم و فعالیت آنزیمی است. سلولهایی که متحمل آپوپتوز میشوند نیز معمولا وزیکولهای خارج سلولی با قطر 5-1 میلیمتر رها میکنند که به آنها اجسام آپوپتوزی میگویند (Dignat-George & Boulanger 2011, van der Pol et al. 2012, Colombo et al. 2014) (تصویر 1).
در برخی مقالات، میکرووزیکولهای جدا شده بوسیلهی سانتریفوژ با عنوان میکرو ذرات شناخته میشوند، مخصوصا آنهایی که از پلاکتها یا سلولهای اندوتلیال منتشر شدهاند. در این مقاله برای وضوح بیشتر وزیکولهای خارج سلولی را بر اساس مکانیسم تولید سلولی و محدودهی سایزشان در دستههای میکرووزیکولها و اگزوزومها قرار میدهیم (Thery et al. 2009)، این رویکرد توسط محققین دیگر نیز مورد استفاده قرار گرفته است، البته این نکته را در نظر بگیرید که روشهای جداسازی قادر به جداسازی یک جمعیت خالص از وزیکولها نیستند. لازم به ذکر است که محدودهی اندازهی وزیکولهای خارج سلولی میتواند همپوشانی داشته باشد و به صورت ویژه، سایز میکرووزیکولها میتواند همپوشانی بالایی با اگزوزومها داشته باشد. در جایی که ترکیبی از اگزوزومها و میکرووزیکولها وجود داشته باشد (مانند زمانی که وزیکولهای پلاسما با سرعت g 100000 سانتریفوژ میشوند)، ما آنها را وزیکول خارج سلولی خواهیم نامید. این وزیکولهای خارج سلولی گاهی اوقات با عنوان وزیکولهای شبه اگزوزومی نیز شناخته میشوند.
یکی از شاخصهای تمام وزیکولهای خارج سلولی حضور فسفاتیدیل سرین (PS) بر روی سطح بیرونی بدلیل فقدان عدم تقارن غشاء در طول ترکیدن (اجسام آپوپتوزی) یا جوانه زنی (میکرووزیکولها) و پیچش به سمت داخل غشاء در طول تشکیل وزیکول در MVBها (اگزوزومها) است. این نکته را میتوان با اتصال انکسین V نشاندار دریافت، معرفی که غالبا برای آنالیز فلوسیتومتری سلولهای آپوپتوزی مورد استفاده قرار میگیرد. با این حال، اخیرا گروههای متعددی توانستهاند میکرووزیکولهای فاقد PS بر روی سطح بیرونی غشاء را شناسایی کنند، این یافته نشان میدهد که فسفاتیدیل سرین برای تشکیل میکرووزیکولها ضروری نیست (Larson et al. 2012, Hou et al. 2014).
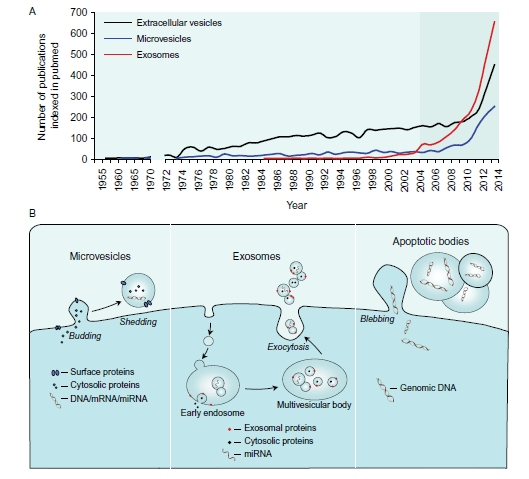
اگزوزومها و میکرووزیکولها هر دو حاوی محمولههایی هستند که آنها را به سلولهای دورتر منتقل خواهند کرد. این محموله میتواند شامل مواد ژنتیکی مانند mRNA، میکرو RNA (miRNA)، یا حتی مقادیر کمی DNA و پروتئینهایی مانند فاکتورهای رونویسی، سیتوکینها و فاکتورهای رشد باشد. به طرز مهمی میکرووزیکولها گیرندههای سلولی و پروتئینهای داخل غشائی را بر روی سطحشان حمل میکنند که مشابه با سلولی است که از آن منشاء گرفتهاند. این ویژگی به شناسایی آنها کمک میکند، اما همچنین بدان معنی است که میتوانند با سلولهای هدف خاصی برهمکنش کرده و آبشار پیامرسانی را از طریق برهمکنش با گیرنده فعال کنند (تداخل پیام رسانی مشابه با برهمکنشهای سلول به سلول) و همچنین اختصاصیت تحویل محموله را افزایش خواهند داد. از سوی دیگر، اگزوزومها با شاخصهایی مانند Alix، HSP70 و تترااسپانین CD9 و CD63 آراسته شدهاند که میتوانند با اتصال به اینتگرین بتا-2 و ارتباط داخل سلولی مرتبط باشند. هرچند این مولکولها معمولا به عنوان شاخصهای اگزوزومی مورد استفاده قرار میگیرند، اما مختص اگزوزومها نیستند و مشخص شده که در وزیکولهای خارج سلولی دیگر نیز وجود دارند. علاوه بر این، تمام وزیکولهای خارج سلولی CD63 را بیان نمیکنند و زیر جمعیتهایی از اگزوزومها میتوانند شاخصهای متفاوتی را بیان کنند (Thery et al. 2009). لازم به ذکر است که اگزوزومها لزوما پروتئینهای شاخص مشابه با سلولهای والدینی را بیان نمیکنند. به عنوان مثال، ما دریافتهایم که شاخص اندوتلیال معمول CD144 بر روی اگزوزومهای منتشر شده از سلولهای اندوتلیال سیاهرگ ناف انسان (HUVECs) وجود ندارند (تصویر 2). تحقیقات اخیر توانستهاند بیان شاخص سطحی اگزوزومها و وزیکولهای خارج سلولی پلاسما را با استفاده از پروفایلگیری گستردهی آنتیبادی تعریف کنند و این نتایج نشان دادهاند که اگزوزومها میتوانند شاخصهای غشائی سطحی مانند CD146، CD4، CD3 و CD45 را بیان کنند (Jorgensen et al. 2015). شواهدی وجود دارد که نشان میدهند محتویات پروتئینی و RNA اگزوزومها به سلول منشاء بستگی دارند (de Jong et al. 2012).
مکانیسم مسئول تشکیل اگزوزومها و بستهبندی انتخابی پروتئینها، لیپیدها و RNA کاملا شناخته نشده، اما به تدریج در حال مشخص شدن است. به نظر نمیرسد که مسیر کمپلکس مرتبساز اندوزومی مورد نیاز برای انتقال برای تولید زیستی اگزوزومها مورد نیاز باشد، اما برخی از اجزاء این مسیر بویژه Alix در تشکیل آنها نقش دارند (Trajkovic et al. 2008, Baietti et al. 2012, Raposo & Stoorvogel 2013). مولکولهای دیگر مانند تتراسپانینها و سرامید که در اگزوزومها به میزان زیاد یافت میشوند نیز در تولید اگزوزومها نقش دارند. بعنوان مثال، مهارکنندههای اسفنگومیلیناز خنثی (یک آنزیم درگیر در تولید سرامید) سبب مهار تشکیل اگزوزومها خواهند شد (Trajkovic et al. 2008). مکانیسم انتشار اگزوزومها نیز به خوبی شناخته نشده است. اعضای خاص خانوادهی Rab GTPase برای انتشار مناسب اگزوزومها لازم هستند، هر چند که اعضای درگیر به نوع سلول و طرح آزمایش بستگی دارند و ممکن است زیرگونههای مختلف اگزوزومهای مرتبط با مرحلهی تشکیل اندوزوم/MVB (اولیه یا ثانویه) را منعکس کنند (Colombo et al. 2014).
ABSTRACT Microvesicles and exosomes: new players in metabolic and cardiovascular disease
The past decade has witnessed an exponential increase in the number of publications referring to extracellular vesicles (EVs). For many years considered to be extracellular debris, EVs are now seen as novel mediators of endocrine signalling via cell-to-cell communication. With the capability of transferring proteins and nucleic acids from one cell to another, they have become an attractive focus of research for different pathological settings and are now regarded as both mediators and biomarkers of disease including cardio-metabolic disease. They also offer therapeutic potential as signalling agents capable of targeting tissues or cells with specific peptides or miRNAs. In this review, we focus on the role that microvesicles (MVs) and exosomes, the two most studied classes of EV, have in diabetes, cardiovascular disease, endothelial dysfunction, coagulopathies, and polycystic ovary syndrome. We also provide an overview of current developments in MV/exosome isolation techniques from plasma and other fluids, comparing different available commercial and non-commercial methods. We describe different techniques for their optical/biochemical characterization and quantitation. We also review the signalling pathways that exosomes and MVs activate in target cells and provide some insight into their use as biomarkers or potential therapeutic agents. In summary, we give an updated focus on the role that these exciting novel nanoparticles offer for the endocrine community.
Introduction
It is well established that patients with metabolic diseases, in particular insulin resistance and type 2 diabetes mellitus (T2DM), are more than twice as likely to develop accelerated cardiovascular disease (CVD) including atherosclerosis, stroke, and coronary artery disease (reviewed in Rask-Madsen & King (2013)). Coronary artery disease is a major cause of morbidity and mortality worldwide, and is a leading cause of death in T2DM, with excess risk of fatality in women compared with men (Peters et al. 2014).
Extensive coronary artery disease can result in myocardial infarction, severe loss of cardiac function, and subsequently lead to the development of heart failure (Hausenloy & Yellon 2013). A cluster of risk factors have recently been defined by the American Diabetes Association and the American College of Cardiology Foundation as reliable indicators of a patient’s risk for T2DM and CVD, and has been defined as cardiometabolic risk (CMR; Brunzell et al. 2008). These risks include obesity, hyperglycemia, hypertension, insulin resistance, and dyslipidemia. The presence of secondary CVD in patients with ischaemia-reperfusion (IR) or T2DM may be referred to as cardio-metabolic disease (CMD). Given its increasing prevalence and severe consequences, new approaches are needed to diagnose and treat CMD. Extracellular vesicles (EVs) are small (50 nm to 2 mm) vesicles released from the surface of many different cell types into different bodily fluids, including plasma, milk, saliva, sweat, tears, semen, and urine. There are several classes of EV, including exosomes, microvesicles (MVs), and apoptotic bodies, which are produced by different mechanisms. Attracting perhaps the most attention recently have been exosomes (50–100 nm), a homogenous population of EV which are released from cells when multivesicular bodies (MVB; sometimes called multivesicular endosomes, MVE) fuse with the plasma membrane in a highly regulated process and release their contents. Cells can also produce a more heterogeneous population of EVs up to 2 mm in diameter called MVs, which are formed by budding and shedding of the cell membrane, a process that involves calcium dependent signalling and enzyme activity. Cells undergoing apoptosis also typically release EV of 1–5 mm in diameter which are referred to as apoptotic bodies (Dignat-George & Boulanger 2011, van der Pol et al. 2012, Colombo et al. 2014) (Fig. 1).
In some literature, MVs isolated by centrifugation are referred to as ‘microparticles,’ particularly those isolated from platelets or endothelial cells. For clarity, this review will refer to EVs simply as exosomes or MV on the basis of the mechanism of their cellular production and their size range – an approach that has been taken by others (Thery et al. 2009), with the caveat that most isolation methods do not provide a pure populations of vesicles. It is important to note that the size ranges of EVs may overlap and in particular, the size of MVs could overlap with the exosomal size range. Where a mixture of exosomes and MV is likely, for example when plasma vesicles are isolated by high speed (w100 000 g) ultracentrifugation, we refer to them more broadly as EV. These EV are sometimes also referred to as ‘exosome-like vesicles’.
One of the characteristic markers of all EVs is the presence on the outer surface of phosphatidylserine (PS), due to loss of membrane asymmetry during blebbing (apoptotic bodies) or budding (MV) and inward folding of the membrane during vesicle formation in MVBs (exosomes). This can be identified by binding of labelled annexin V, a reagent often used for flow cytometric analysis of apoptotic cells. However, more recently several groups have identified MVs lacking PS on the outer membrane, suggesting that this is not essential for MV formation (Larson et al. 2012, Hou et al. 2014).
Both exosomes and MVs characteristically carry a cargo, which they are able to deliver to cells in remote locations. The cargo can include genetic material such as mRNA, microRNA (miRNA), or even small amounts of DNA (Moldovan et al. 2013), and proteins including transcription factors, cytokines, and growth factors, have also been described. Importantly, MVs also carry cellular receptors and transmembrane proteins on their surface characteristic of the cells from which they were released. This aids in their identification but also means that they can interact with specific target cells instigating signalling cascades via receptor interactions (rececrine signalling – akin to cell–cell interactions) and also increasing specificity of cargo delivery. On the other hand, exosomes are characteristically decorated with markers including Alix, HSP70, and the tetraspanins CD9 and CD63, which may be associated with beta-2 integrin binding and intercellular communication. Although these are commonly used as markers of exosomes, they are not exclusive to exosomes and may be found on other EVs. Furthermore, not all EVs express CD63 and different sub-populations of exosomes may express different markers (Thery et al. 2009). It is important to consider that exosomes do not necessarily express the same marker proteins as their parent cells. For example, we found that the common endothelial marker CD144 is absent on exosomes from human umbilical vein endothelial cells (HUVECs) (Fig. 2). Recent work has further defined plasma EV and exosome surface marker expression by using extensive antibody profiling which showed that exosomes can express surface membrane markers such as CD146, CD4, CD3, and CD45 (Jorgensen et al. 2015). There is some evidence that the protein and RNA content of exosomes depends on the state of the source cell (de Jong et al. 2012).
The mechanism behind the formation of exosomes and selective packaging of proteins, lipids, and RNA is not completely understood but is gradually becoming revealed. The Endosomal Sorting Complex Responsible for Transport pathway does not seem to be required for exosome biogenesis, although some components are involved in their formation, particularly Alix (Trajkovic et al. 2008, Baietti et al. 2012, Raposo & Stoorvogel 2013). Other molecules that are enriched in exosomes such as tetraspanins and ceramide have also been implicated in exosome biogenesis. For example, inhibitors of neutral sphingomyelinase, an enzyme involved in ceramide production, inhibits exosome production (Trajkovic et al. 2008). Less well understood is the mechanism of exosome release. Certain members of the Rab GTPase family are required for efficient release of exosomes, although the exact members involved appears to depend on the cell type and experimental design, and may reflect different subtypes of exosomes relating to the stage (early or late) of endosome/MVB formation (Colombo et al. 2014).
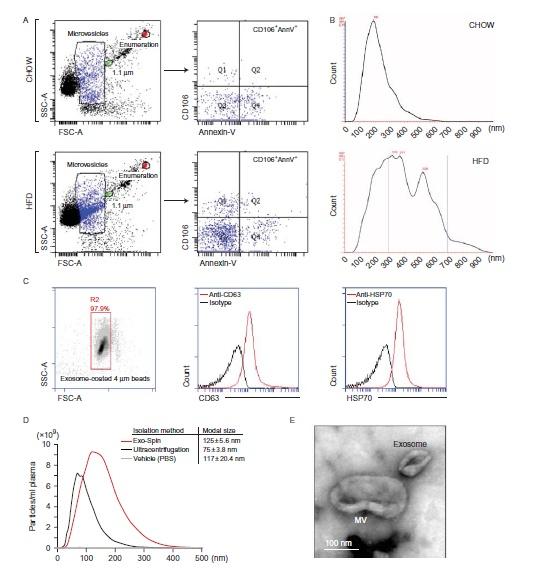
- مقاله درمورد نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- نقش میکرووزیکولها و اگزوزومها در دیابت، بیماریهای قلبی عروقی، اختلالات اندوتلیال، کواگولوپاتی و سندروم تخمدان پلیکیستی
- Microvesicles و exosomes: بازیکنان جدید در بیماری های متابولیکی و قلب و عروق
- پروژه دانشجویی نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- نقش میکرووزیکول و اگزوزوم ها در بیماری های متابولیک و قلبی و عروقی
- پایان نامه در مورد نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- تحقیق درباره نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- مقاله دانشجویی نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق در قالب پاياننامه
- پروپوزال در مورد نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- گزارش سمینار در مورد نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق
- گزارش کارورزی درباره نقش میکرووزیکول ها (MVs) و اگزوزوم ها در بیماری های متابولیکی و قلب و عروق