بررسی و مطالعه دیانای متیلاسیون در حیوانات

14,500 تومانشناسه فایل: 5013
1- مقدمه
توالی متیله غالب در تمام حیوانات، دی نوکلئوتید CpG خود مکمل است. در مهره داران، اکثر CpGs ها در ژنوم، در موقعیت 5 روی حلقه سیتوزین، متیله شده قرار دارند. چندین پیامد بیولوژیکی این تغییر یا اصلاح پس سنتتیک، شناخته شده است. یکی از بهترین این پیامدها، جهش زایی مرتبط با متیلاسیون است که سبب می شود CpG در ژنوم ایجاد شود و مسئول یک سوم جهش های نقطه ای است که منجر به افزایش بیماری های ژنتیکی در انسان ها می شوند (Bird, 1980; Jones et al., 1992).
دیدن این امر به عنوان یک مزیت انتخابی متیلاسیون DNA ، دشوار است. احتمالا به صورت یک بهاء غیرقابل اجتناب برای برخی مزایای دیگر CpG-متیل، پرداخت می شود. در بی مهرگان (که بیش از 95% گونه های حیوانی را تشکیل می دهند)، حتی اگر هر کدام از ژن های آنها متیله شود، ممکن نیست که این بها را بپردازند. با بررسی اختلال در ژن متیل ترانسفراز سیتوزین (MTase) در موش ها، لزوم متیلاسیون DNA در طول رشد و نمو طبیعی پستانداران، نشان داده شده است(Li et al., 1992).
جهش در مرحله جنینی تا حد زیادی سطح متیلاسیون DNA را کاهش می دهد، و در اواسط بارداری، جنین می میرد. به دنبال این بحث، برای توضیحی در مورد فنوتیپ کشنده جنینی، سعی شده است که اثرات متیلاسیون بر روی رونویسی را مورد بررسی قرار دهند. متیلاسیون DNA، با سرکوب رونویسی مرتبط است. که این سرکوب، توسط وارد کردن ساختارهای متیله مصنوعی به درون سلولها (Vardimin et al., 1982; Stein et al., 1982), و با استفاده از داروهایی که MTase را مهار می کنند، نشان داده شده است (Jones and Taylor, 1980).
فرضیه منطقی و معقول به این صورت است که جنینی که فاقد MTase باشد، می میرد، زیرا مکانیسم سرکوب با واسطه متیلاسیون آن، با شکست مواجه می شود.
ABSTRACT Studies of DNA methylation in animals
We have been studying the evolution and function of DNA methylation in vertebrate animals using three related approaches. The first is to further characterise proteins that bind to methylated DNA. Such proteins can be viewed as ‘receptors’ of the methyl-CpG ‘ligand’ that mediate downstream consequences of DNA modification. The second approach involves CpG islands. These patches of non-methylated DNA coincide with most gene promoters, but their origin and functional significance have only recently become the subject of intensive study. The third approach is to trace the evolution of DNA methylation. Genomic methylation patterns of vertebrates are strikingly different from those of invertebrates. By studying methylation in animals that diverged from common ancestors near to the invertebrate/vertebrate boundary, we will assess the possibility that changes in DNA methylation contributed causally to the evolution of the complex vertebrate lineage.
INTRODUCTION
The predominant methylated sequence in all animals is the self-complementary dinucleotide CpG. In vertebrates, most CpGs in the genome are methylated at the 5 position on the cytosine ring. Several biological consequences of this postsynthetic modification are known. Best understood is the methylation-associated mutagenesis that has caused the under-representation of CpG in the genome and is responsible for over one third of the point mutations that give rise to human genetic diseases (Bird, 1980; Jones et al., 1992). It is difficult to see this as a selected advantage of DNA méthylation. More likely it is an unavoidable price to be paid for some other benefit of methyl-CpG. Strikingly, the invertebrates (which account for well over 95% of animal species) may not pay this price, as few, if any of their genes are methylated (see below). The need for DNA méthylation during normal mammalian development has been shown by disruption of the gene for cytosine methyltransferase (MTase) in mice (Li et al., 1992).
Mutant embryos have greatly reduced levels of DNA méthylation, and die in mid-gestation. In seeking an explanation for this embryonic lethal phenotype, it is tempting to focus on the well-known effects of méthylation on transcription. DNA méthylation has long been correlated with transcriptional repression. That it causes repression has been shown by introduction of artificially methylated constructs into cells (Vardimin et al., 1982; Stein et al., 1982), and by the use of drugs that inhibit the MTase (Jones and Taylor, 1980). A reasonable hypothesis is that embryos lacking the MTase die because the methylation-mediated repression mechanism fails.
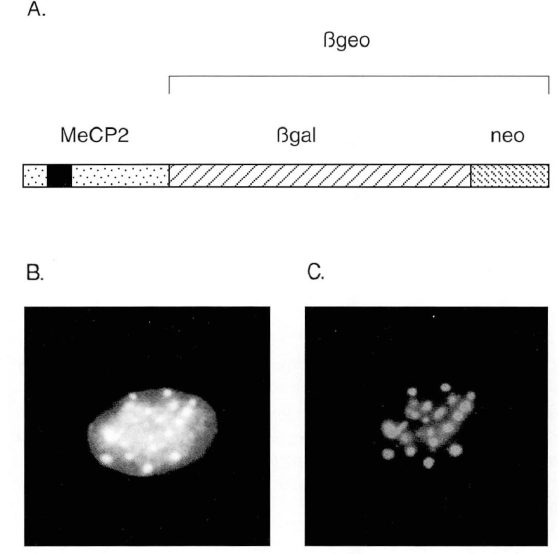
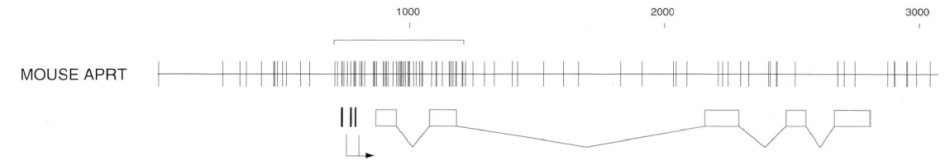
- مقاله درمورد بررسی و مطالعه دیانای متیلاسیون در حیوانات
- پروژه دانشجویی بررسی و مطالعه دیانای متیلاسیون در حیوانات
- متیلاسیون دی ان ای در حیوانات
- پایان نامه در مورد بررسی و مطالعه دیانای متیلاسیون در حیوانات
- تحقیق درباره بررسی و مطالعه دیانای متیلاسیون در حیوانات
- مقاله دانشجویی بررسی و مطالعه دیانای متیلاسیون در حیوانات
- بررسی و مطالعه دیانای متیلاسیون در حیوانات در قالب پاياننامه
- پروپوزال در مورد بررسی و مطالعه دیانای متیلاسیون در حیوانات
- گزارش سمینار در مورد بررسی و مطالعه دیانای متیلاسیون در حیوانات
- گزارش کارورزی درباره بررسی و مطالعه دیانای متیلاسیون در حیوانات