نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور

شاپا پرینت: 1674-800X وبسایت مرجع 48 رفرنس دارد
13,500 تومانشناسه فایل: 10688
مقدمه مقاله
پروتئین های ROR1 و ROR2 پروتئین غشای تراوا خانواده کیناز تیروزین گیرنده (RTK) هستند. البته ROR1 / 2 در ابتدا در یک لاین سلولی نوروبلاستوما در یک غربالگری PCR برای کیناز تیروزین گیرنده کشف شد و سابقا به عنوان تیروزین کیناز نوروتروفیک مربوط به گیرنده (NTRKR1 / 2) نامگذاری می شد. این ROR1/2 انسانی دارای 58٪ خصوصیت اسید آمینه ای است و به طور نزدیکی CLO با گیرنده های خانواده MUSK و TRK مرتبط هستند (مسیاکوسکای و کارول، 1992؛ فورستر و همکاران، 1999).
هر دو ژن پروتئین هایی با وزن مولکولی 104 کیلو دالتون پیش بینی شده را رمز گذاری کردند، اما ROR1 دارای چند سایت Nگلیکوزیلاسیون است که از نظر ترجمه ای تولید پست ROR1 در 130 کیلو دالتون می کند. این سایت Nگلیکوزیلاسیون رفت و آمد ROR1 به غشا و به نوبه خود عملکرد ROR1 مهم است (کائوکا و همکاران، 2011).
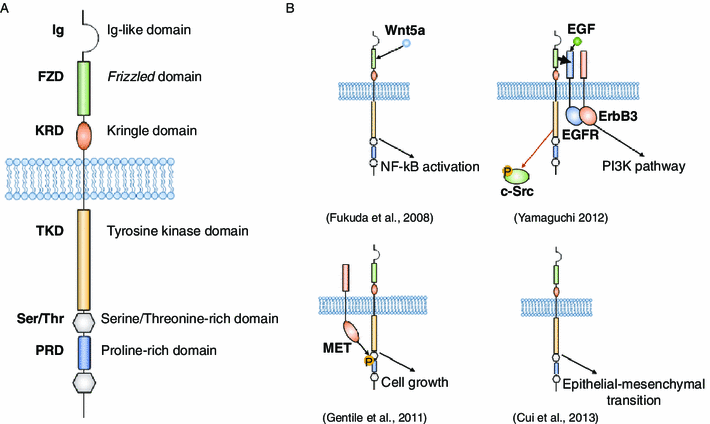
ساختار ROR1/2 شامل دامنه ایمونوگلوبولین مانند (IG) خارج سلولی در انتهای آمینه می باشد، که پیرو یک دامنه غنی از سیستئین شناخته شده به عنوان یک دامنه مجعد (FZD)، و سپس یک دامنه Kringle (KRD) در یک دامنه غشای تراوا است (شکل A1). دامنه FZD در گیرنده های خانواده صاف و مجعد، همچنین کربوکسی پپتیداز Z ، کلاژن هجدهم α1، و پروتئین های مربوط به گیرنده لیپوپروتئین با چگالی کم (LRP) دیده شده و شامل 10 باقی مانده سیستئین حفظ شده و 5 پیوند دی سولفید مربوطه می باشد.
این FZD تعامل گیرنده لیگاند واسطه تصور می شود (روزموسز و همکاران، 2001؛ فورستر و همکاران، 2004؛ . میکلز و نوسه، 2006). هم ROR1 که به میزان بیشتری در مقالات بررسی شده، و هم ROR2، نشان دهنده پیوند Wnt5a،Wnt غیر متعارف از طریق FZD است (ایشی و همکاران، 2003؛ میکلز و نوسه، 2006؛ فوکودا و همکاران، 2008؛ پاگانونی و همکاران، 2010). این KRD بسیار چین خورده، دامنه غنی از سیستئین که واسطه ها در تعامل پروتئین – پروتئین و پروتئین – لیگاند در پروتئین انعقادی، آپولیپوپروتئینها و فاکتور رشد سلول های کبدی می باشد (استفانز و همکاران 1992، میزونو و همکاران، 1994؛. متیوز همکاران، 1996).
بخش سیتوپلاسمی انسانی ROR1 / 2 دارای یک دامنه تیروزین کیناز (TKD)است، و منتج از یک دامنه غنی سرین / ترئونین (SER / THR)، یک دامنه غنی از پرولین (PRD)، و یک دامنه Ser/Thr دوم در انتهای کربوکسی است (شکل A1). عاملیت دامنه تیروزین کیناز (TKD) از RORs در نوشتارها مورد بحث قرار گرفته است. مطالعات اولیه نشان دهنده فعالیت قوی کیناز اوتوکاتالیتیک برای ROR2 است، در حالی که نشان دهنده ROR1 ضعیف برای فعالیت کیناز متوسط است (ماسیاکواسکای و کارول، 1992؛ ایشی و همکاران، 1999).اخیرا، ROR1 TKD دیده شده برای فسفریلات C-SRC در سلول های NIH3T3 کافی بود (یاماگوچی و همکاران، 2012). در مقابل، یکی دیگر از مطالعه نتیجه گرفت بیایند که ROR1 کیاناز دورغین به صورت ROR1 بوده که هیچ فعالیت کینازی را در سلول های COS-7 را نشان نمی دهد (جنتیل و همکاران، 2011). خانواده های ROR یکی از متفاوت ترین خانواده ها در خانواده کیناز تیروزین گیرنده محسوب می گردد و فقط حاوی 21 از 40 باقی مانده مشترک در دیگر TKDs شرح داده شده توسط هنکس و کوین می باشد (هنکس و همکاران، 1988). قابل ذکر است،ROR1 دارای زیر نهادهایی در C482G، K614R و L634Fبود، که باید پیوند ATP و عملکرد کیناز را تعدیل می کرد (هنکس و همکاران 1988،؛ ماسیاکواسکای و کارول، 1992).خانواده ROR از پروتئین هایی هستند از روی قواعد تکامل حفظ شده و سطح بالایی از شباهت بین orthologs درMus musculu ،elgans Caenorhabditis ، Xenopus laevevis ، Drosophila melnogaster،Apylasia californica، و Gallus gallus را به اشتراک گذاشته بود (ویلسون و همکاران، 1993؛. فورستر و همکاران .، 1999؛ ایشی و همکاران، 1999؛ مککی و همکاران، 2001؛ هیکاسا و همکاران، 2002؛ استیکر و همکاران، 2006).
اینCAM-1 ROR1 / 2 ortholog در الگانس C منحصر به فرد است یک مشخصه اسید آمینه بیشتری را برای ROR1 به اشتراک گذاشته، اما فاقد PRD و دامنه Ser/Thr دوم است. البته DROR یک واسطه ساختاری از خانواده گیرنده ROR و TRK است و DRNK Drophosilia orthologs هستند و فاقد دامنه Ig خارج سلولی و PRD داخل سلولی و دامنه های Ser/Thr می باشد (ویلسون و همکاران 1993، ایشی و همکاران، 1997). حفاظت RORs در میان گونه های مختلف اهمیت خانواده ROR را از طریق تعدادی از فرآیندهای در طول تکامل مشخص می کند.
ABSTRACT ROR1, an embryonic protein with an emerging role in cancer biology
Receptor tyrosine kinase-like orphan receptor 1 (ROR1) is a member of the ROR family consisting of ROR1 and ROR2. RORs contain two distinct extracellular cysteine-rich domains and one transmembrane domain. Within the intracellular portion, ROR1 possesses a tyrosine kinase domain, two serine/threonine-rich domains and a proline-rich domain. RORs have been studied in the context of embryonic patterning and neurogenesis through a variety of homologs. These physiologic functions are dichotomous based on the requirement of the kinase domain. A growing literature has established ROR1 as a marker for cancer, such as in CLL and other blood malignancies. In addition, ROR1 is critically involved in progression of a number of blood and solid malignancies. ROR1 has been shown to inhibit apoptosis, potentiate EGFR signaling, and induce epithelial-mesenchymal transition (EMT). Importantly, ROR1 is only detectable in embryonic tissue and generally absent in adult tissue, making the protein an ideal drug target for cancer therapy.
Introduction
ROR1 and ROR2 are transmembrane proteins within the receptor tyrosine kinase (RTK) family. ROR1/2 were initially discovered in a neuroblastoma cell line in a PCR screen for receptor tyrosine kinases and were formerly named as neurotrophic tyrosine kinase receptor-related (NTRKR1/2). Human ROR1/2 have 58% amino acid identity and are closely related to MUSK and Trk family receptors (Masiakowski and Carroll, 1992; Forrester et al., 1999).
Both genes encode proteins with a predicted molecular weight of 104 kDa, but ROR1 has multiple N-glycosylation sites that generate post-translationally modified ROR1 at 130 kDa. These N-glycosylation sites are necessary for the trafficking of ROR1 to the membrane and in turn the function of ROR1 (Kaucká et al., 2011). The structure of human ROR1/2 consists of an extracellular immunoglobulin-like (Ig) domain at the amino-terminus, followed by a cysteine-rich domain known as a Frizzled domain (FZD), and then a Kringle domain (KRD) into a transmembrane domain (Fig. 1A).
The FZD domain is seen in the Smoothened and Frizzled-family receptors, as well as carboxypeptidase Z, collagen α1 XVIII, and low-density lipoprotein receptor-related proteins (LRP) and consists of 10 conserved cysteine residues and five corresponding disulfide bonds. The FZD is thought to mediate receptor-ligand interaction (Roszmusz et al., 2001; Forrester et al., 2004; Mikels and Nusse, 2006). Both ROR1 and to a greater extent in the literature, ROR2, have been shown to bind Wnt5a, a non-canonical Wnt via the FZD (Oishi et al., 2003; Mikels and Nusse, 2006; Fukuda et al., 2008; Paganoni et al., 2010). The KRD is a highly-folded, cysteine-rich domain that mediates in protein-protein and protein-ligand interaction in coagulation proteins, apolipoproteins, and hepatocyte growth factor (Stephens et al., 1992; Mizuno et al., 1994; Mathews et al., 1996).
The cytoplasmic portion of human ROR1/2 has a tyrosine kinase domain (TKD), followed by a Serine/Threonine-rich domain (Ser/Thr), a proline-rich domain (PRD), and a second Ser/Thr domain at the carboxy-terminus (Fig. 1A). The functionality of the tyrosine kinase domain (TKD) of RORs has been debated in the literature. Early studies showed strong autocatalytic kinase activity for ROR2, while ROR1 possessed weak to moderate kinase activity (Masiakowski and Carroll, 1992; Oishi et al., 1999).
More recently, ROR1 TKD was shown to be sufficient to phosphorylate c-SRC in NIH3T3 cells (Yamaguchi et al., 2012). Conversely, another study concluded that ROR1 is a pseudokinase as ROR1 did not show any kinase activity in COS-7 cells (Gentile et al., 2011). The ROR families are one of the most divergent within the receptor tyrosine kinase family, containing only 21 of the 40 consensus residues in other TKDs described by Hanks and Quinn (Hanks et al., 1988). Notably, ROR1 possesses substitutions at C482G, K614R and L634F, that should modulate ATP binding and kinase function (Hanks et al., 1988; Masiakowski and Carroll, 1992).
The ROR family of proteins are evolutionally conserved and share a high level of homology between orthologs in Mus musculus, Caenorhabditis elgans, Xenopus laevevis, Drosophila melnogaster, Apylasia californica, and Gallus gallus (Wilson et al., 1993; Forrester et al., 1999; Oishi et al., 1999; McKay et al., 2001; Hikasa et al., 2002; Stricker et al., 2006). CAM-1 is the singular ROR1/2 ortholog in C. elegans and shares a greater amino acid identity to ROR1, but lacks the PRD and the second Ser/Thr domain. DROR, a structural intermediate of the ROR and TRK receptor family, and DRNK are the Drophosilia orthologs and lack the extracellular Ig domain and the intracellular PRD and Ser/Thr domains (Wilson et al., 1993; Oishi et al., 1997). The conservation of RORs across species underlies the importance of the ROR family through a number of processes during evolution.
- مقاله درمورد نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- ROR1، یک پروتیین جنینی با نقش تازه در زیست شناسی سرطان
- پروژه دانشجویی نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- نقش پروتئین جنینی ROR1 در سرطان
- پایان نامه در مورد نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- تحقیق درباره نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- مقاله دانشجویی نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور در قالب پاياننامه
- پروپوزال در مورد نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- گزارش سمینار در مورد نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور
- گزارش کارورزی درباره نقش پروتئین جنینی ROR1 در زیست شناسی سرطان با یک نقش نوظهور